中鎖脂肪ケトン食療法とレチノイドなどを併用した「がん細胞の分化誘導と増殖抑制」療法
【エピジェネティクスとは】
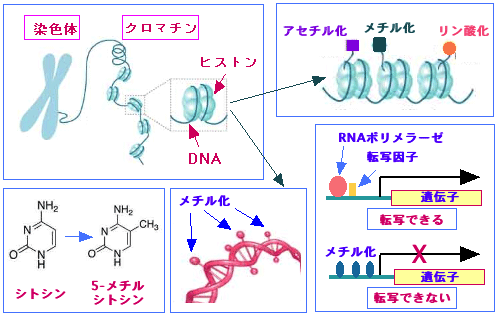
細胞の遺伝情報は細胞核の中の遺伝子(=DNA)に入っています。人間の1個の細胞の核には約30億対のヌクレオチドからなるDNA(デオキシリボ核酸)が格納されており、このDNAが遺伝子の本体です。ヒト遺伝子は約22000種類のタンパク質を作る設計図(遺伝情報)をもっていますが、このゲノムの持つ遺伝情報の発現は、DNAの塩基配列と転写装置だけで制御されているわけではなく、DNAのメチル化やヒストン(DNAに結合するタンパク質)の修飾などによって複雑に制御されています。
「ゲノム(遺伝子)の塩基配列以外のメカニズムで遺伝子発現を制御し、細胞や生体に変化を生じさせる現象」のことを、エピジェネティクス(epigenetics)と言います。エピジェネティクスの「エピ」はギリシャ語の接頭語で「上にある、別の、後から」という意味で、本来の遺伝情報(DNAの塩基配列)の土台の「上にかぶさる別の遺伝情報」や「後天的に獲得した遺伝情報」という意味を示しています。
例えば、私たちの体を構成する細胞は全て同じ遺伝情報を持っていますが、皮膚や神経や筋肉や肝臓など機能の異なる細胞になれるのは、それぞれの細胞において、使う遺伝子と使わない遺伝子に違いがあるからです。約22000個の遺伝子の全てが発現しているわけではなく、発現している遺伝子の違いによって細胞の種類が決まります。このように、DNAの塩基配列(=遺伝情報)が同じなのに、使う遺伝子と使わない遺伝子に目印をつけて、細胞に変化を生じさせる現象がエピジェネティクスです。細胞のがん化の主な原因は、遺伝子の突然変異によるがん遺伝子やがん抑制遺伝子の機能異常によると考えられています。しかし、最近の研究によって、遺伝子の変異とは関係ない、エピジェネティック(epigenetic)な機序によるがん遺伝子やがん抑制遺伝子の発現異常による発がんメカニズムの重要性が指摘されるようになりました。
遺伝子にはその発現を調節する部分があり、これをプロモーターと言います。遺伝子を使うか使わないかを制御している領域のことです。遺伝子が発現するためには、DNAからメッセンジャーRNAを作るRNAポリメラーゼという酵素や遺伝子発現を調節する転写因子がこのプロモーター領域に結合することが必要です。このプロモーター領域には、CpG(C はシトシン、Gはグアニン)という配列が繰り返された部分があり、DNAメチル化とは、DNAのCpGという配列の部分でC(シトシン)にメチル基(-CH3)いう分子がつくことです。プロモーター領域のDNAにメチル化が起こると、RNAポリメラーゼや転写因子が結合できなくなり、遺伝子からmRNAが転写される段階が阻害され、遺伝子発現のスイッチがオフになります。このように、エピジェネティスによって遺伝子発現のスイッチが切られることを「遺伝子のサイレンシング(silencing)」と呼ばれています。
遺伝子サイレンシングの別の方法にヒストンの修飾があります。例えば、ヒストンのアセチル化はDNAとヒストンの結合を緩くすることによって遺伝子の転写を促進します。
図:DNAのメチル化やヒストンの修飾(アセチル化など)によって遺伝子の発現が変化することをエピジェネティクスと言う。
【ヒストンのアセチル化とは】
細胞核内では、DNAはヒストンという球状の蛋白質複合体に巻き付くような状態で存在します。
ヒストンはリシン(リジン)やアルギニンといった塩基性(プラスの電荷をもつ)のアミノ酸が多く、酸性(マイナスの電荷をもつ)のDNAと強い親和性を持っています。ヒストンは、長いDNAをコンパクトに核内に収納するための役割と同時に、遺伝子発現の調節にも重要な役割を果たしています。
ヒストンによる遺伝子発現の調節は複雑ですが、簡単にまとめると、「ヒストンとDNAの結合は転写に阻害的に働く」ということです。遺伝子が転写されるためには、転写因子やRNAポリメラーゼなどの他の蛋白質がDNAに結合する必要があり、ヒストンが結合していると転写に邪魔になります。したがって、転写の活発な遺伝子の部分ではヒストンとDNAの結合が緩くなっています。
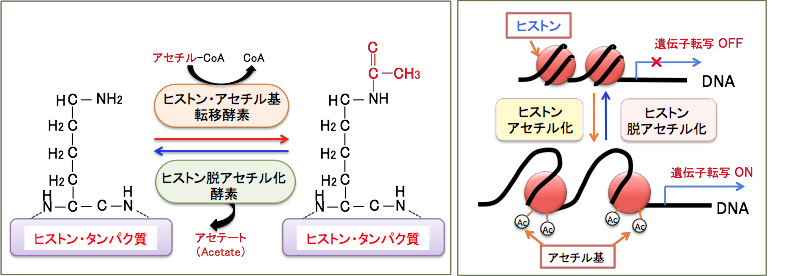
DNAとヒストンの結合を緩くする機序として、「ヒストンのアセチル化」という現象があります。アセチル化というのはアセチル(CH3CO)基が結合することです。ヒストンのN末端領域のリシン残基のアミノ基(-NH2)がアセチル化という修飾を受けるとアミド(-NHCOCH3)に変換し、リシン残基の塩基性が低下して酸性のDNAとの親和性が無くなり、DNAからヒストンが離れ、DNAが露出することになります。
一般的に、ヒストンが高度にアセチル化されている領域の遺伝子は転写が活発に行われていることを示しています。すなわち、ヒストンのアセチル化は遺伝子発現を促進(正に制御)し、 反対に、ヒストンが脱アセチル化(低アセチル化)されることにより遺伝子発現は抑制(負に制御)されると考えられています。
ヒストンのアセチル化と脱アセチル化の反応は「ヒストンアセチル基転移酵素(=ヒストンアセチルトランスフェラーゼ)」と「ヒストン脱アセチル化酵素(=ヒストンデアセチラーゼ)」によってダイナミックに制御されており、遺伝子発現のON/OFFのメインスイッチになっていると考えられています(下図)。
【βヒドロキシ酪酸はヒストン脱アセチル化酵素を阻害して、ヒストンのアセチル化を亢進する】
βヒドロキシ酪酸はケトン体の一種です。ケトン体は絶食などで糖質が枯渇した状態で脂肪酸の燃焼(β酸化)が亢進したときに肝臓で産生され、グルコース(ブドウ糖)が枯渇した時の代替エネルギーになります。絶食時などで日常的に産生されています。
ケトン体としてアセト酢酸、βヒドロキシ酪酸、アセトンの3種が作られますが、アセトンは呼気となって排泄され、アセト酢酸とβヒドロキシ酪酸はエネルギー源になります。
最近の論文で、「βヒドロキシ酪酸はヒストン脱アセチル化酵素を阻害し、酸化ストレス耐性遺伝子の発現を亢進させ、酸化ストレスを抑制する」ことが示されています。(Science 339(6116): 211-4, 2013年)
この論文で以下のような結果が得られています。
1)培養細胞やマウスの動物実験でβヒドロキシ酪酸はクラスIヒストン脱アセチル化酵素を阻害する。
2)飢餓や直接βヒドロキシ酪酸を投与する方法でマウスの血中のβヒドロキシ酪酸の濃度(0.6~1.5mM)を上昇させると、腎臓など複数の臓器においてヒストンのアセチル化が増えていることが確認できた。
3)βヒドロキシ酪酸によって発現が亢進する遺伝子として、細胞周期の停止や酸化ストレス耐性にかかわる転写因子をコードしているFOXO3a遺伝子や、酸化ストレス耐性遺伝子のMt2が同定された。つまり、βヒドロキシ酪酸はクラスIヒストン脱アセチル化酵素を阻害することによってFOXO3aやMt2などの酸化ストレス耐性遺伝子のプロモーターにおけるヒストンの高アセチル化を引き起こしていると考えられた。
4)酸化ストレス耐性に重要は働きを果たしているMnスーパーオキシドジスムターゼおよびカタラーゼは転写因子FOXO3aの標的遺伝子である。βヒドロキシ酪酸の投与によって、マウスの腎臓において、FOXO3aのみならず、Mnスーパーオキシドジスムターゼとカタラーゼの発現量もタンパク質レベルで顕著に増加していた。
5)除草剤のパラコートをマウスに投与して酸化障害によるタンパク質のカルボニル化を測定すると、βヒドロキシ酪酸を投与されたマウスでは、酸化ストレスによって引き起こされるカルボニル化を顕著に抑制した。以上の結果から、βヒドロキシ酪酸は内在性のヒストン脱アセチル化酵素を阻害し、酸化ストレスに対して抵抗性を高める遺伝子のプロモーター部分におけるヒストンの高アセチル化を引き起こし、これらの遺伝子発現を亢進することによって酸化ストレスを軽減するということが証明されたということです。
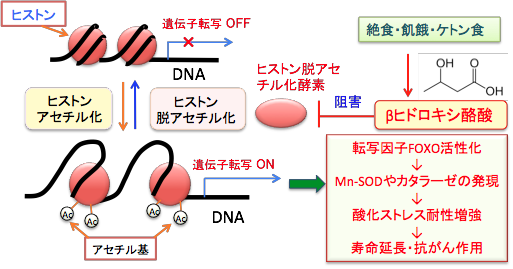
酸化ストレスに対する抵抗性の亢進は寿命延長やがん抑制に有効であることが明らかになっています。さらに、ヒストン脱アセチル化酵素の阻害が、寿命の延長やがん細胞の分化誘導や増殖抑制に効果があることも多くの研究で明らかになっています。つまり、絶食やケトン食による健康作用や寿命延長作用や抗がん作用の一部は、βヒドロキシ酪酸によるヒストン脱アセチル化酵素の阻害作用が関与している可能性を示唆しています。
ケトン食は成長ホルモン/インスリン様成長因子-1のシグナル伝達系に抑制的に作用することが明らかになっています。さらに、ヒストンのアセチル化による寿命延長と抗がん作用も加わります。糖質を制限し、脂肪酸の燃焼を増やし、ケトン体を多く出させる中鎖脂肪ケトン食が、美容と寿命延長と抗がん作用を同時に達成できることを示す科学的エビデンスが増えています。
図:絶食や飢餓やケトン食によってβヒドロキシ酪酸の濃度が上昇すると、ヒストン脱アセチル化酵素が阻害される。これによって遺伝子プロモーター領域のヒストンが高アセチル化し、転写因子FOXOが活性化され、Mnスーパーオキシドジスムターゼ(Mn-SOD)やカタラーゼの発現を誘導し、酸化ストレスに対して耐性が増強する。
【レチノイドは遺伝子の発現を調節する】
レチノイド(retinoid)はビタミンA(レチノール)およびその誘導体や類縁化合物の総称です。レチノイドは生体内では活性型であるAll-trans retinoic acid (ATRA:以下レチノイン酸)として細胞核内の受容体に結合して、その生理作用を発揮します。
レチノイン酸の核内受容体には、レチノイン酸受容体(retinoic acid receptor: RAR)とレチノイドX受容体(retinoid X receptor: RXR)があり、それぞれα、β、γのサブタイプが存在します。
これらの受容体はリガンドの結合刺激によりホモ二量体(RAR-RARやRXR-RXR)を形成しますが、RXRはRXRとのホモ二量体だけでなく、RARやビタミンD受容体などの核内受容体とのヘテロ二量体(RAR-RXR)も形成します。
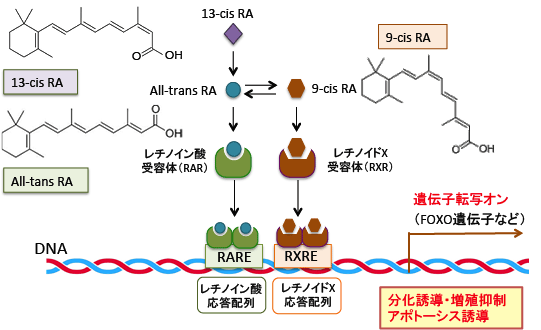
そして、これらの二量体は標的遺伝子のプロモーター領域に存在するレチノイン酸応答配列(retinoic acid response element: RARE)あるいはレチノイドX応答配列(retinoid X response element: RXRE)と結合することによって、様々な標的遺伝子の発現を調節しています。
異性体の関係にあるAll-trans-RA (ATRA) と 9-cis-RA (9C-RA)は2つともRARのリガンドになりますが、RXRのリガンドとなるのは9C-RAのみです。
レチノイドによって発現が調節される遺伝子は細胞の分化や増殖や死(アポトーシス)の制御に重要は働きを担っているため、その機能異常は細胞のがん化に関連し、ある種のがんに対してレチノイドが効く場合があります。
例えば、レチノイン酸の二重結合がすべてトランス型になったall-trans retinoic acid(ATRA)は急性前骨髄球性白血病の特効薬になっています。また、RXRαのアゴニストとして合成された非環式レチノイドが肝臓がんの再発を予防する効果が報告されています。ある種のがんにはレチノイドは増殖抑制や分化誘導やアポトーシス誘導などの抗がん作用を示します。
難治性にきびの治療に用いられている13-cisレチノイン酸(イソトレチノイン)は、にきびに対する作用機序において、IGF-1産生抑制作用や転写因子のFOXOを活性化する作用などが報告され、また、単独では抗腫瘍効果は弱いのですが、他の治療との組合せで抗腫瘍効果が高まることが報告されています。13-cisレチノイン酸は体内でAll-trans レチノイン酸に変換されて効果を発揮します。
図:ビタミンAの体内における活性本体であるAll-trans retinoic acid(レチノイン酸)はレチノイン酸受容体(retinoic acid receptor: RAR)に結合するとホモ二量体を形成し、標的遺伝子のプロモーター領域にあるレチノイン酸応答配列(RARE)に結合して遺伝子転写のスイッチをオンにする。レチノイン酸の異性体である9-cis retinoic acidはレチノイン酸受容体(RAR)の他にレチノイドX受容体(RXR)にも結合して、ホモ二量体やヘテロ二量体を形成して細胞核内の受容体に結合して標的遺伝子の転写を誘導して生理機能を発揮する。13-cis retinoic acid(イソトレチノイン)はプロドラッグであり、細胞内でAll-trans RAに変換されて、同様の遺伝子発現の調節を行う。
【イソトレチノイン(13-cisレチノイン酸)はIGF-1の産生を低下させる】
成長期の子供がイソトレチノイン(13-cis retinoic acid)を服用すると、成長が止まることが報告されています。その理由として成長ホルモンやインスリン様成長因子-1への作用が指摘されています。イソトレチノインがインスリン様成長因子-1(IGF-1)の血中濃度を低下させる作用が報告されています。
イソトレチノインは小児の腫瘍の神経芽細胞腫(neuroblastoma)の治療に効果があります。この神経芽細胞腫の治療に13-cisレチノイン酸を使った小児の成長が抑制されたという報告があります。(Endocr J. 46 Suppl:S113-5, 1999年)
例えば、47人のにきび患者(平均年齢21.5±5.1歳)を対象に、イソトレチノインの1日量を最初の1ヶ月間は0.5〜0.75mg/kgで開始し、その後維持量として0.88 mg/kgで治療を行っています。3ヶ月後に採血して検討したところ、インスリン様成長因子-1とインスリン様成長因子結合蛋白-3の血中濃度が著明に低下していました。(Br J Dermatol 162(4): 798-802, 2010年)
イソトレチノインはIGF-1シグナル伝達系を抑制的に作用するようです。
【イソトレチノイン(13-cisレチノイン酸)は転写因子FOXOの発現を亢進する】
転写因子FOXO(Forkhead Box O)はDNA結合ドメインFOX(Forkhead box)を持つForkheadファミリーのサブグループ“O”に属する転写因子です。哺乳類ではFOXO1,3a,4,6の4種類が存在します。FoxO1とFoxO3は約650個のアミノ酸からなる蛋白質で、遺伝子のプロモーター領域のTTGTTTACという配列に結合 します。アンドロゲン受容体やβカテニンとも相互作用します。
転写因子というのは特定の遺伝子の発現(DNAの情報を蛋白質に変換すること)を調節している蛋白質で、FOXOはストレス応答、代謝制御、細胞周期、アポトーシス、細胞分化、DNA修復、免疫機能、炎症などに関連する多くの遺伝子の発現を促します。FOXOは様々なストレスに対する抵抗力を高める作用を担っており、たとえば、カタラーゼやスーパーオキシドディスムターゼ(SOD)のような抗酸化酵素の発現を高めることによって酸化ストレスに対する抵抗力を高める作用があります。
がん抑制遺伝子としての性格ももっており、FOXOの活性化は抗がん作用を示します。
FOXO はインスリン/PI3K/Aktシグナルによって負に制御されます。FOXOは細胞質と核内をシャトルすることのよって活性が調節されています。 フォスフォイノシトール-3-キナーゼ(PI3K)によってリン酸化されて活性化されたセリン・スレオニン・キナーゼのAktは、細胞核内に入ってFOXOをリン酸化します。FOXOはAktによってリン酸化されると核の外(細胞質)に移行します。したがって、PI3K/Aktシグナル伝達系を活性化するインスリンやインスリン様成長因子-1はFOXOの転写活性を抑制することになるのです。
栄養飢餓状態では、PI3K/Aktの不活化によってFOXOの転写活性が高まります。つまり、FOXOはカロリー制限における寿命延長や老化性疾患の抑制において重要な役割を果たしていることが知られています。
イソトレチノインがFOXO1とFOXO3aの活性を高めることが報告されています(イソトレチノインはFOXO3aの活性を高め、FOXO3aがFOXO1の転写を亢進します)。(G Ital Dermatol Venereol. 145(5):559-71. 2010年)
にきび(医学用語では尋常性痤瘡)の発生には、乳製品や高糖質食でインスリン/IGF-1シグナル伝達系が活性化され、mTORC1の活性化やFOXO1の核外移行が促進されることが関連しています。FOXO1の転写活性が低下すると、男性ホルモン受容体の発現が増し、皮脂腺の分泌が増えます。イソトレチノインはFOXO1の発現を亢進し、AMPKを活性化し、mTORC1の活性を抑制します。つまり、イソトレチノインはIGF-1の産生を低下させ、さらにFOXO1を介してmTORC1の活性を低下させるので、にきびを治す効果があるというわけです。
以上のことをまとめたのが以下の図です。
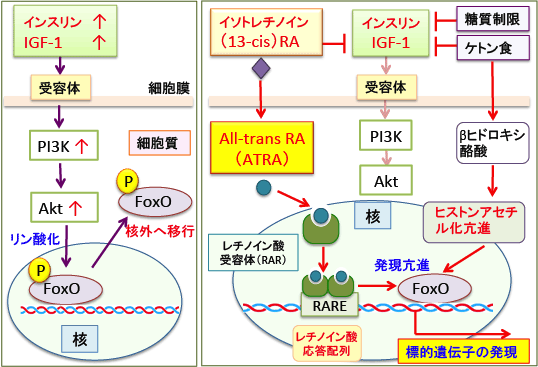
図:(左)インスリン/インスリン様成長因子-1(IGF-1)はPI3K/Aktシグナル伝達系を亢進し、活性化されたAktは転写因子FoxOをリン酸化する。リン酸化されたFoxOは核外(細胞質)へ移行するので、FoxOの転写活性は抑制される。
(右)イソトレチノイン(13-cis retinoic acid)は細胞内でAll-transレチノイン酸
(ATRA)に変換され、レチノイン酸受容体に結合し二量体を形成して標的遺伝子のプロモーター部分のレチノイン酸応答配列(RARE)に結合して遺伝子発現を亢進する。イソトレチノインはIGF-1の産生を抑制する作用がある。さらに、糖質制限はインスリン濃度を低下させ、ケトン食はIGF-1濃度を低下させると同時に、ヒストン脱アセチル化酵素を阻害してヒストンのアセチル化を亢進してFoxO遺伝子の発現を亢進する作用もある。したがって、ケトン体を多く産生する中鎖脂肪ケトン食とイソトレチノインは転写因子のFoxOの活性化などによって相乗効果で抗がん作用を強めることができる。
【ヒストンのアセチル化促進作用とイソトレチノインの相乗効果】
13-cisレチノイン酸(イソトレチノイン)は体内でall-transレチノイン酸(ATRA)に変換され、ATRAは細胞核内のレチノイン酸受容体(retinoic acid receptor:RAR)やレチノイドX受容体(retinoid X receptor: RXR)に結合して2量体を形成して、標的遺伝子のプロモーター領域に存在するレチノイン酸応答配列(retinoic acid response element: RARE)あるいはレチノイドX応答配列(retinoid X response element: RXRE)と結合することによって、様々な標的遺伝子の発現を調節しています。
にきびの治療にはイソトレチノインは劇的に効きます。がんに対する効果も期待されて多くの臨床試験が行われたのですが、あまり良い結果は得られなかったため、次第に注目されなくなっています。
ただ、がん細胞の遺伝子にはエピジェネティックは制御が起こっています。特に細胞増殖を抑制したり細胞死や分化を誘導するようながん抑制遺伝子には、DNAのアセチル化やヒストンの脱アセチル化によって転写因子がアクセスできにくくなっていることが明らかになっています。つまり、レチノイドが思うように効かないのは、これらの遺伝子にレチノイドがアクセスできないためで、アクセスできるようすればレチノイドの抗がん作用が出てくる可能性があります。
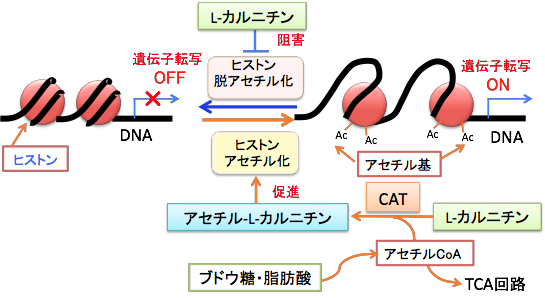
中鎖脂肪ケトン食は、糖質と極力摂取しないので、インスリン/インスリン様成長因子-1(IGF-1)シグナル伝達系を抑制し、ケトン体のβヒドロキシ酪酸は、ヒストン脱アセチル化酵素を阻害することによって、ヒストンの高アセチル化によってFOXO遺伝子などのがん抑制作用のある遺伝子の発現を促進します。ヒストンのアセチル化によって転写因子がアクセスしやすくなると、レチノイドの遺伝子発現誘導作用も促進されます。つまり、にきび治療薬の13-cisレチノイン酸(イソトレチノイン)を併用するとFOXO遺伝子の発現と転写活性を高めることができます。さらに、脂肪酸の燃焼を促進してケトン食の効果を高めるL-カルニチンにはヒストンのアセチル化を高めて、がん細胞の増殖を抑制することが示されています。すなわち、L-カルニチンは内因性のヒストン脱アセチル化酵素の阻害剤で、生体内でがん細胞の増殖を選択的に阻害することが報告されていま。(PLoS One. 2012; 7(11): e49062.)
さらに、L-カルニチンにアセチル基(CH3CO-)が結合したアセチル-L-カルニチンはヒストンアセチル化のアセチル基を供給する作用があります。
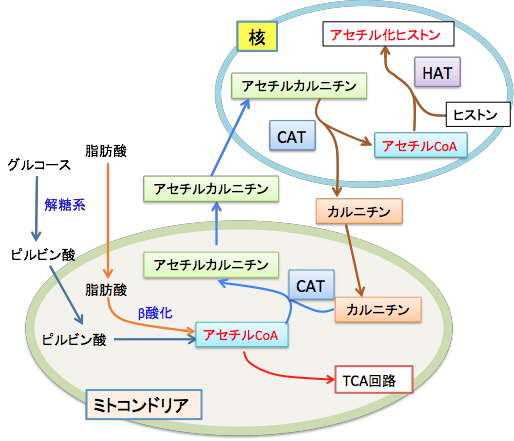
ミトコンドリアで生成されたアセチルCoAは、カルニチン・アシルカルニチン・トランスロカーゼ(carnitine acylcarnitine translocase)の作用で細胞質に運ばれ、ついで細胞核に運ばれ、核でカルニチン・アセチルトランスフェラーゼ(carnitine acetyltransferase)によってアセチルCoAに変換され、ヒストンのアセチル化のアセチル基の供給源となることが報告されています。(Epigenetics 4(6):399-403, 2009年)アセチル-CoAはグルコース(ブドウ糖)や脂肪酸の分解で生成されます。すなわち、グルコースが解糖系で代謝されてピルピン酸が作られ、ピルビン酸がミトコンドリアに入って、ピルビン酸脱水素酵素の働きでアセチル-CoAに変換されてTCA回路に入ります。脂肪酸もミトコンドリアでβ酸化によって分解されてアセチル-CoAに変換されTCA回路に入ります。このとき、グルコースが枯渇しているとアセチル-CoAは肝臓でケトン体生成に使われます。
グルコース枯渇時にアセチル-CoAがケトン体に変換されるのは、アセチル-CoAが細胞膜を通れないので、細胞膜を通過できるケトン体に変換されて脳などの他の組織の細胞にエネルギー産生の原料として運ばれるためです。
細胞核におけるヒストンのアセチル化では、アセチル-CoAのアセチル基が使われますが、このアセチル-CoAはミトコンドリアで作成され、ミトコンドリアから核への運搬にはL-カルニチンが必要ということです。この経路をまとめると以下のようになります。
図:グルコースや脂肪酸の分解によって産生されたミトコンドリア内のアセチル-CoAは、カルニチンアセチルトランスフェラーゼ(CAT)の作用により、L-カルニチンと結合してアセチル-L-カルニチンとしてミトコンドリア外の細胞質に輸送され、さらに細胞核に移行する。核に移行したアセチル-L-カルニチンは、L-カルニチンとアセチル-CoAに分解され、アセチル-CoAはヒストンアセチルトランスフェラーゼ(HAT)の作用によってヒストンをアセチル化する。
図:L-カルニチンにはヒストン脱アセチル化酵素阻害作用によってヒストンのアセチル化のレベルを高める。さらにL-カルニチンはアセチル-L-カルニチンとしてミトコンドリアで生成されたアセチルCoAを核に運ぶ役割も担っている。核ではアセチルCoAのアセチル基をヒストンアセチル基転移酵素の働きでヒストンをアセチル化する。つまり、L-カルニチンとアセチル-L-カルニチンは、ヒストンのアセチル化を促進する。
緑茶に含まれるエピガロ・カテキン・ガレート(EGCG)が、がん抑制遺伝子のDNAのメチル化を阻害してがん抑制遺伝子の働きを高めることが報告されています。EGCGのがん予防効果は今まで主に抗酸化作用によるものと考えられてきましたが、DNAメチル化の阻害でがん抑制遺伝子の活性を高める作用も関与しているかもしれないということです。その他、ウコンのクルクミンや、赤ぶどうの皮に含まれるレスベラトロール、アブラナ科野菜のスルフォラファン、大豆のゲニステインなど、がん予防効果が報告されている様々な成分についても、DNAメチル化やヒストンのアセチル化の調節などエピジェネティクスに作用する結果が報告されています。
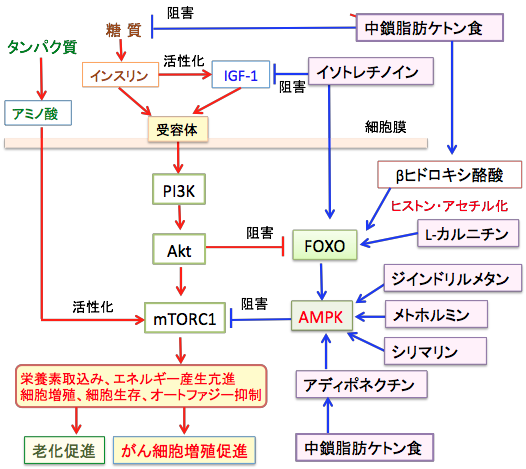
さらに、AMP依存性プロテインキナーゼ(AMPK)を活性化してmTORC1(哺乳類ラパマイシン標的蛋白複合体-1)メトホルミン、シリマリン、ジインドリルメタンなどの併用は、抗がん作用を強化できます。
図:中鎖脂肪ケトン食は糖質摂取が少ないため、インスリン/インスリン様成長因子-1(IGF-1)シグナル伝達系を抑制する。ケトン体のβヒドロキシ酪酸とL-カルニチンはヒストンの高アセチル化を引き起こし、転写因子のFOXOの発現を亢進する。レチノイドのイソトレチノインはIGF-1の産生抑制とFOXO発現を亢進する。
ケトン食はアディポネクチンの産生を高めてAMPKの活性を亢進する。メトホルミン、ジインドリルメタン、シリマリンもAMPKを活性化する。AMPKの活性化はmTORC1の活性を阻害することによってがん細胞の増殖を抑制する。これらの相乗効果で抗がん作用を強めることができる。
●1ヶ月分の費用の目安 :
イソトレチノイン(Tretiva):3000円(1日10mg)〜 12000円(1日40mg) |
|
L-カルニチン:6300〜12600円 |
メトホルミン:3000〜9000円 |
シリマリン:5000円 |
ジインドリルメタン:9000円 |